

Мышечно-суставные рецепторы
Необходимым условием нормальной мышечной деятельности является получение информации о положении тела в пространстве и о степени сокращения каждой из мышц. Эта информация поступает в центральную нервную систему от рецепторов вестибулярного аппарата, глаз, кожи, а также от проприорецепторов (мышечно-суставных рецепторов). К проприорецепторам относятся:
- мышечные веретена, находящиеся среди мышечных волокон,
- тельца Гольджи, расположенные в сухожилиях,
- пачиниевы тельца, находящиеся в фасциях, покрывающих мышцы, в сухожилиях, связках и периосте.
Все эти проприорецепторы относятся к группе механорецепторов. Мышечные веретена и тельца Гольджи возбуждаются при растяжении, а пачиниевы тельца — при давлении.
О важности афферентной иннервации мышц свидетельствует и тот факт, что 30-50% волокон в любом нерве, подходящем к мышце, являются афферентными, несущими импульсацию от проприорецепторов. К уже отмечалось, при выключении афферентной иннервации мышц, например при перерезке задних корешков спинного мозга, координированная мышечная деятельность нарушается, несмотря на сохранение двигательной иннервации мышц.
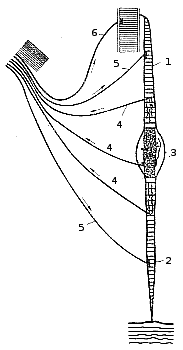 |
Мышечные веретена представляют собой высокодифференцированные рецепторные образования, снабженные афферентными и эфферентными нервными волокнами. Каждое веретено состоит из нескольких тонких так называемых интрафузальных поперечно-полосатых мышечных волокон (рис. 195). Oдиночное волокно состоит из центральной части — ядерной сумки и двух способных к сокращению участков. В ядерной сумке pacположены рецепторы, представляющие сообой спиралевидные окончания толстых афферентных покрытых миелиновой оболочкой нервных волокон. Сокращающиеся участки интрафузального волокна иннервированы тонкими моторными, так называемыми гамма-эфферентными, волокнами, образующими мелкие концевые пластинки. Рис. 195. Схематическое изображение мышечного веретена кролика. 1 — проксимальный конец интрафузального мышечного волокна, прикрепленный к волокну скелетной мышцы; 2 — дистальный конец интрафузального волокна,прикрепленный к фасции; 3 — ядерная сумка; 4 — афферентные волокна, идущие от рецепторов интрафузального волокна; 5 — гамма-эфферентные волокна; 6 — моторное волокно, идущее к скелетной мышце. |
Один конец интрафузального волокна прикрепляется к обычному мышечному волокну, а другой — к сухожилию. Таким образом, интрафузальные волокна расположены параллельно массе мышечных волокон, среди которых они находятся. Если мышца растянута или расслаблена, то мышечные веретена тоже растянуты и в рецепторах ядерной сумки возникают импульсы, идущие в центральную нервную систему. Если же мышца сокращена, то натяжение мышечных веретен ослабевает и импульсация прекращается.
Второй вид проприорецепторов — тельца Гольджи — находится в сухожилиях. При мышечном сокращении тельца Гольджи растягиваются и в них возникают нервные импульсы, поступающие к нервным центрам. Сухожильные рецепторы менее возбудимы, чем мышечные веретена.
Таким образом, в двигательном аппарате имеются рецепторы — мышечные веретена,— возбуждающиеся при удлинении (расслаблении и растяжении), и рецепторы — тельца Гольджи,— возбуждающиеся при сокращении мышечных волокон.
Импульсы, поступающие в центральную нервную систему от мышечных веретен, облегчают возникновение рефлекторной реакции данной мышцы и тормозят сокращение мышцы-антагониста. Импульсы, приходящие от сухожильных рецепторов — телец Гольджи, вызывают противоположные рефлекторные реакции (рис. 180).
Импульсация из мышечных веретен, как показал Р. Гранит, может изменяться с помощью специального регуляторного механизма. Такой механизм образован сократительными элементами, расположенными по обе стороны от ядерной сумки интрафузального волокна. Их сокращение вызывает растяжение ядерной сумки и раздражение рецепторов веретена. Таким образом, эти рецепторы могут посылать в центральную нервную систему интенсивную импульсацию даже при небольшом сокращении или расслаблении поперечнополосатой мышцы.
Степень сокращения сократительных элементов веретена регулируется гамма-эфферентными нервными волокнами, которые являются отростками, гамма-мотонейронов спинного мозга. Импульсы, приходящие по гамма-эфферентным волокнам, вызывают сокращения интрафузальных мышечных волокон, что приводит к усилению потока афферентных импульсов от растягиваемых рецепторов ядерной сумки. Активность гамма-мотонейронов регулируется ретикулярной формацией среднего мозга .
Сократительные элементы мышечных веретен всегда находятся в состоянии некоторого тонуса, так как по гамма-эфферентам непрерывно поступают к ним импульсы из центральной нервной системы. Это влечет собой афферентную импульсацию от мышечных веретен, которая в свою ередь тонизирует мотонейроны спинного мозга, что является одной из причин рефлекторного тонуса скелетной мускулатуры. Отличительная особенность проприорецепторов — их малая способность к адаптации, благодаря чему центральная нервная система получает непрерывные сигналы о состоянии скелетной мускулатуры и может осуществлять непрерывную регуляцию двигательных актов.
Кинестетические сигналы, т. е. сигналы о движении отдельной части тела, играют чрезвычайно важную роль в развитии восприятий организма. Они являются главным контролем других органов чувств, например, зрения. Так, оценка зрением расстояния до какого-нибудь предмета вырабатывается при помощи мышечного чувства. Возбуждения, связанные с работой мышц, возникающие при хватании предмета руками или приближении к нему, сочетаются со зрительными ощущениями и служат важнейшим критерием для выработки суждения о том, насколько отдален предмет.
