

На протяжении долгого времени пользовалась широким признанием резонаторная теория Г. Гельмгольца (1863), объяснявшая механизм восприятия звуков различной высоты (разной частоты колебаний). Гельмгольц обратил внимание на то, что идущие в поперечном направлении плотные волокна, образующие основную перепонку имеют разную длину. У основания улитки они наиболее коротки (их длина на 0,04 мм); у ее вершины они значительно длиннее (0,5 мм). Гельмгольц предполагал, что волокна основной перепонки натянуты подобно струнам арфы и имеют разную собственную частоту колебаний, на которую они способны резонировать. При действии звука в наибольшей степени начинает колебаться то волокно, которое «настроено» в резонанс данной частоте колебаний, т. е. способно ее воспроизводить. Колебание каждого из волокон основной мембраны воспринимается рецепторными клетками кортиева органа, расположенными именно на этом волокне. Таким образом, по Гельмгольцу, для тона каждой высоты существует свой индивидуальный рецептор.
Ряд фактов заставил отказаться от теории Гельмгольца. Прежде всего не подтвердилось предположение, что волокна основной мембраны находятся в натянутом состоянии, подобно струнам арфы. Далее установлено, что при действии тонов определенной высоты возникают колебания одного какого-либо волокна основной мембраны, а широкого ее участка. Кроме того, при звучании низких тонов импульсы возникают в болшом количестве волокон слухового нерва, а не в отдельных его волокнах, как это следовало предполагать по теории Гельмгольца.
Другой теорией слуха была телефонная теория Резерфорда (1880), которая предполагала, что частоты колебаний потенциалов в слуховом нерве при восприятии звуков разной высоты соответствуют частотам принимаемых звуков, как это происходит в телефонном кабеле при передаче звуков по телефону. После открытия микрофонного эффекта улитки казалось, что он подтверждает телефонную теорию. Однако вскоре выяснилось, что частоты импульсов в слуховом нерве при действии высоких звуков (выше 1000 колебании в секунду) не соответствуют частоте звуковых колебаний. Так как рефрактерный период даже быстро проводящих нервных волокон не бывает ниже 1 мсек, то, следовательно, информация о действующих на ухо высоких тонах не может передаваться в центральную нервную систему в виде синхронных со звуком колебаний потенциала, т. е. по принципу, используемому в телефоне. Для передачи информации о звуковых колебаниях высокой частоты необходимо ее преобразование.
В настоящее время можно считать выясненным вопрос о механизме восприятия звуков разной высоты. Некоторые представления, легшие в основу и резонаторной и телефонной теории, оказались справедливыми.
Как было сказано выше, звуковые колебания высокой частоты вовлекают в колебательный процесс перилимфу верхнего и нижнего каналов улитки, т. е. вестибулярной и барабанной лестницы, не на всем протяжении этих каналов. Воспринимаемые мембраной овального окна колебания вызывают колебательные движения столба жидкости (перилимфы и эндолимфы) лишь на ограниченном участке вблизи овального окна. Колебания перилимфы верхнего канала передаются на перилимфу нижнего канала через основную мембрану на определенном расстоянии от начала улитки. Это расстояние тем меньше и, следовательно, место передачи колебаний с верхнего канала на нижний расположено тем ближе к овальному и круглому окну, чем выше тон, действующий на улитку.
В улитке имеют место явления резонанса. Однако резонирующим субстратом является не определенное волокно основной перепонки, как предполагал Гельмгольц, а столб жидкости определенной длины. По данным Г. Бекеши, чем выше звук, т. е. чем больше частота воспринимаемых ухом колебаний, тем меньше длина колеблющегося столба жидкости в каналах улитки и тем ближе к основанию улитки и овальному окну расположено место максимальной амплитуды колебания.
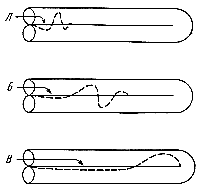 |
При действии звуков низкой частоты длина колеблющегося столба жидкости увеличивается и место максимальной амплитуды колебания отодвигается в сторону вершины улитки (рис. 202). При колебаниях жидкости в каналах улитки колеблется и основная мембрана, причем не отдельные волокна ее, а большие или меньшие (по длине) ее участки. Естественно, что при этом будет возбуждено разное количество рецепторных клеток, расположенных на этой мембране. Рис. 202. Схема колебаний основной мембраны при высоких (А), средних (Б) и низких (В) звуках. |
При действии звуков низкой частоты окажутся возбужденными репепторные клетки вдоль всей основной мембраны; при действии же высоких тонов будет возбуждаться меньшее число клеток — только те, которые расположены на основной мембране основания улитки.
Для восприятия звуков разной высоты имеет существенное значение локализация рецепторных клеток кортиева органа, вовлекаемых в возбуждение. Это доказывается многими физиологическими экспериментами. Л. А. Андреев в лаборатории И. П. Павлова выработал у собаки условные рефлексы на тоны различной высоты, затем разрушал кортиев орган одного уха и повреждал тонкой иглой разные участки кортиева органа второго уха. Если был поврежден первый завиток улитки у ее основания, то исчезали условные рефлексы на высокие тона; если же повреждение было нанесено в области вершины улитки, то исчезали условные рефлексы на низкие тона; разрушение только среднего завитка улитки вызывало падение тонов среднего диапазона.
В опытах на морских свинках, улитка которых имеет 4½ завитка, Г. Девис обнаружил, что микрофонный эффект при отведении электрических потенциалов от основания улитки наблюдается при всех звуковых частотах, а при отведении от участков ее, расположенных ближе к ее вершине, только при действии низких звуков.
Приведенные факты приводят к заключению, что при действии тонов разной высоты, т. е. при действии звуковых колебаний разной частоты, в улитке происходит пространственное кодирование звуковой информации. Каждой высоте тона соответствует определенная длина участка основной мембраны, охваченного колебательным процессом, и определенное место на этой мембране, через которое колебания перилимфы верхнего канала передаются на перелимфу нижнего. Вследствие этого каждой высоте ктона соответствует определенное количество рецепторов, в которых возникает процесс возбуждения, а также определенная группа рецепторов, сила раздражения которых оказывается наибольшей.
При звуковых частотах выше 100—150 колебаний в секунду участок максимальной амплитуды колебаний основной мембраны не доходит до ее конца у вершины улитки и передача колебаний перилимфы с верхнего канала на нижний происходит через эту мембрану. При более низких частотах передача колебаний перилимфы происходит через геликотрему, что ведет к раздражению всех слуховых клеток. В таком случае пространственное кодирование становится невозможным. Однако оно в этих случаях становится и ненужным, так как колебания низкой частоты могут без искажений воспроизводиться волокнами слухового нерва. В этом случае информация о высоте тона может передаваться соответствующей частотой импульсов, проходящих по слуховому нерву, подобно тому как проходит передача электрических колебаний звуковой частоты по телефонному кабелю.
Таким образом, существует два механизма различения высоты тонов. При низких тонах информация о них передается по волокнам слухового нерва в виде импульсов, частота которых соответствует частоте звуковых колебаний, воспринимаемых улиткой. При действии же высоких тонов происходит пространственное кодирование.
В определенном диапазоне тонов средней высоты с частотой колебаний до 1000 в секунду (по некоторым данным и несколько более высокой) могут действовать обе системы кодирования звуковой информации: одна, основанная на передаче импульсов, частота которых соответствует частоте раздражения, и вторая, основанная на неодинаковом расположении и одинаковом количестве возбужденных рецепторных клеток.
