

Из венозной крови можно извлечь 55—58 об.% углекислого газа. Большая часть СО2, извлекаемого из крови, происходит из имеющихся в плазме и эритроцитах солей угольной кислоты и только около 2,5 об.% углекислого газа растворено и около 4—5об.% находится в соединении с гемоглобином в виде карбогемоглобина.
Образованно угольной кислоты из углекислого газа происходит в эритроцитах, где содержится фермент карбоангидраза, являющийся мощным катализатором, ускоряющим реакцию гидратации СО2.
Карбоангидраза. Существование этого фермента предполагал еще И. М. Сеченов, но открыт он был лишь в 1932 г. Мелдрумом и Рафтоном.
Связывание углекислого газа кровью в капиллярах большого круга. Углекислый газ, образующийся в тканях, диффундирует в кровь кровеносных капилляров, так как напряжение СО2 в тканях значительно превышает его напряжение в артериальной крови. Растворяющийся в плазме СО2 диффундирует внутрь эритроцита, где под влиянием карбоангидразы он мгновенно превращается в угольную кислоту,
Согласно расчетам, активность карбоангидразы в эритроцитах такова, что реакция гидратации углекислоты ускоряется в 1500—2000 раз. Так как весь углекислый газ внутри эритроцита превращается в угольную кислоту, то напряжение СО2 внутри эритроцита близко к нулю, поэтому все новые и новые количества СО2 поступают внутрь эритроцита. В связи с образованием угольной кислоты из СО3 в эритроците концентрация ионов НСО3' возрастает, и они начинают диффундировать в плазму. Это возможно потому, что поверхностная мембрана эритроцита проницаема для анионов. Для катионов мембрана эритроцита практически непроницаема. Взамен ионов НСО3' в эритроциты входит ион хлора. Переход ионов хлора из плазмы внутрь эритроцита освобождает в плазме ионы натрия, которые связывают поступающие нз эритроцита ионы НСО3, образуя NaHCО3 Химический анализ плазмы венозной крови показывает значительное увеличение в ней бикарбоната.
Накопление внутри эритроцита анионов приводит к повышению осмотического давления внутри эритроцита, а это вызывает переход воды из плазмы через поверхностную мембрану эритроцита. В результате объем эритроцитов в капиллярах большого круга увеличивается. При исследовании с помощью гематокрнта установлено, что эритроциты занимают 40% объема артериальной крови и 40,4% объема венозной крови. Из этого следует, что объем эритроцитов венозной крови больше, чем эритроцитом артериальной, что объясняется проникновением в них воды.
Одновременно с поступлением СО2 внутрь эритроцита и образованием в нем угольной кислоты происходит отдача кислорода оксигемоглобином и превращение его в редуцированный гемоглобин. Последний является значительно менее диссоциирующей кислотой, чем оксигемоглобин и угольная кислота. Поэтому при превращении оксигемоглобина в гемоглобин Н2СО3 вытесняет из гемоглобина ионы калия и, соединяясь с ними, образует калиевую соль бикарбоната.
Освобождающийся Н˙ ион угольной кислоты связывается гемоглобином. Так как редуцированный гемоглобин является малодиссоциированной кислотой, то при этом не происходит закисления крови и разница рН венозной и артериальной крови крайне невелика. Происходящую в эритроцитах тканевых капилляров реакцию можно представить следующим образом:
КНbO2 + Н2СO3= HHb + O2 + КНСO3
Из изложенного следует, что оксигемоглобин, превращаясь в гемоглобин и отдавая связанные им основания углекислоте, способствует образованию бикарбоната и транспорту в таком виде углекислоты. Кроме того, гкмоглобин образует химическое соединение с СО2 — карбогемоглобин. Наличие в крови соединения гемоглобина с углекислым газом было установлено путем следующего опыта. Если к цельной крови прибавить цианистый калий, который полностью инактивирует карбоангидразу, то оказывается, что эритроциты такой крови связывают больше СО2, чем плазма. Отсюда был сделан вывод, что связывание СО2 эритроцитами после инактивирования карбоангидразы объясняется наличием в эритроцитах соединения гемоглобина с СО2. В дальнейшем выяснилось, что СО2 присоединяется к аминной группе гемоглобина, образуя так называемую карбаминовую связь.
Реакция образования карбогемоглобина может идти в одну или другую сторону в зависимости от напряжения углекислого газа в крови. Хотя небольшая часть всего количества углекислого газа, которое может быть извлечено из крови, находится в соединении с гемоглобином (8—10%), однако роль этого соединения в транспорте углекислоты кровью достаточно велика. Примерно 25—30% углекислого газа, поглощаемого кровью в капиллярах большого круга, вступает в соединение с гемоглобином, образуя карбогемоглобин.
Отдача СО2 кровью в легочных капиллярах. Вследствие более низкого парциального давления СО2 в альвеолярном воздухе по сравнению с напряжением его в венозной крови углекислый газ переходит путем диффузии из крови легочных капилляров в альвеолярный воздух. Напряжение СО2 в крови падает.
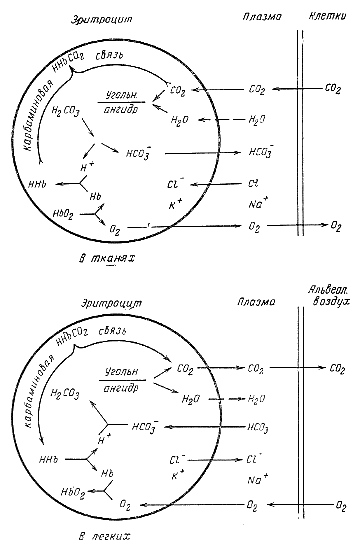 |
Одновременно с этим вследствие более высокого парциального давления кислорода в альвеолярном воздухе по сравнению с его напряжением в венозной крови кислород поступает из альвеолярного воздуха в кровь капилляров легких. Напряжение О2 в крови возрастает, и гемоглобин превращается в оксигемоглобин. Так как последний является кислотой, диссоциация которой значительно выше, чем гемоглобина угольной кислоты, то он вытесняет угольную кислоту из ее калиевой. Реакция идет следующим образом: ННb + O2 + КНСO3= КНbO2+H2CO3 Освободившаяся из своей связи с основаниями угольная кислота расщепляется карбоангидразой на углекислый газ в воду. Значение карбоангидразы в отдаче углекислого газа в легких видно из следующих данных. Для того чтобы произошла реакция дегидратации Н2СО3 растворенной в воде, с образованием того количества углекислого газа, которое выходит из крови за время ее нахождения в капиллярах легких, требуется 300 секунд. Кровь же проходит через капилляры легких в течение 1—2 секунд, но за это время успевает произойти дегидратация угольной кислоты внутри эритроцита и диффузия образовавшегося СО2 сначала в плазму крови, а затем в альвеолярный воздух. Так как в легочных капиллярах уменьшается в эритроцитах концентрация ионов НСО3, то эти ионы из плазмы начинают диффундировать в эритроциты, а ионы хлора диффундируют из эритроцитов в плазму. В связис тем что напряжение углекислого газа в крови легочных капилляров уменьшается, карбаминовая связь расщепляется и карбогемоглобин отдает углекислый газ. Схематически все эти процессы приведены на рис. 57. Рис. 57. Схема процессов, происходящих в эритроците при поглощении или отдаче кровью кислорода и углекислого газа. |
Кривые диссоциации соединений угольной кислоты в крови. Как мы уже говорили, свыше 85% углекислого газа, которое может быть извлечено из крови подкислении ее, освобождается в результате расщепления бикарбонатов (калия в эритроцитах и натрия в плазме).
Связывание углекислого газа и отдача его кровью зависят от его парциального напряжения. Можно построить кривые диссоциации соединений углекислоты в крови, подобные кривым диссоциации оксигемоглобина. Для этого по оси ординат откладывают объемные проценты связанного кровью углекислого газа, а по оси абсцисс— парциальные напряжения углекислого газа. Нижняя кривая на рис. 58 показывает связывание углекислого газа артериальной кровью, гемоглобин которой почти полностью насыщен кислородом. Верхняя кривая показывает связывание кислого газа венозной кровью.
 |
Различие в высоте этих кривых зависит от того, что артериальная кровь, богатая оксигемоглобином, обладает меньшей способностью связывать углекислый газ по сравнению с венозной кровью. Являясь более сильной кислотой, чем угольная кислота, оксигемоглобин отнимает основания у бикарбонатов и этим способствует освобождению угольной кислоты. В тканях оксигемоглобин, переходя в гемоглобин, отдает связанные с ним основания, увеличивая связывание кислого газа кровью. Рис. 58. Кривые поглощения углекислого газа кровью (объяснение в тексте). |
Точка А на нижней кривой на рис. 58 соответствует напряжению кислоты, равному 40 мм рт. ст., т. е. тому напряжению, которое фактически имеется в артериальной крови. При таком напряжении связано 52 об.% СО2. Точка V на верхней кривой соответствует напряжению кислого газа 46 мм рт. ст., т. е. фактически имеющемуся в венозной крови. Как видно из кривой, при таком напряжении венозная кровь связывает 58 об.% углекислого газа. Линия AV, соединяющая верхнюю и нижнюю кривую, соответствует тем изменениям способности связывать углекислый газ, которые происходят при превращении артериальной крови в венозную или, наоборот, венозной крови в артериальную.
Венозная кровь благодаря тому, что содержащийся в ней гемоглобин переходит в оксигемоглобин, в капиллярах легких отдает около 6 об.% СО2. Если бы в легких гемоглобин не превращался в оксигемоглобин, то, как видно из кривой, венозная кровь при имеющемся в альвеолах парциальном давлении углекислого газа, равном 40 мм рт. ст.. связывала бы 54 об.% СО2, следовательно, отдала бы не 6, а только 4об.%. Равным образом, если бы артериальная кровь в капиллярах большого круга не отдавала своего кислорода, т. е. если бы гемоглобин ее оставался насыщенным кислородом, то эта артериальная кровь при парциальпом давлении углекислого газа, имеющемся в капиллярах тканей тела, смогла бы связат не 58 об.% СО2, а лишь 55 об.%.
Таким образом, переход гемоглобина в оксигемоглобин в легких и оксигемоглобина в гемоглобин в тканях тела способствует поглощению и отдаче примерно 3—4 об.% углекислого газа из тех 6 об.%, которые поглощает кровь в тканях и отдает в легких. Около 25—30% выделяемого в легких углекислого газа переносится карбогемоглобином.
Из всего сказанного вытекает, что в механизме транспорта и кислорода, и углекислого газа кровью важнейшая роль принадлежит эритроцитам, в которых содержатся гемоглобин и карбоангидраза.
