

Электрические явления в сетчатке и зрительном нерве
Фотохимические изменения зрительных пигментов палочек и колбочек представляют собой начальное звено в цепи явлений возбуждения зрительных рецепторов при световом раздражении. Вслед за комплексом фотохимических реакций в зрительных рецепторах, а затем и в зрительном нерве возникают электрические колебания, связанные с возбуждением сложного рецепторного аппарата глаза.
Электрические колебания, носящие название электроретинограммы могут быть обнаружены при световом раздражении и отведении разности электрических потенциалов от неповрежденного глаза или же непосредственно от сетчатки.
Для записи электроретинограммы в эксперименте на вырезанном глазе один электрод прикладывают к роговой оболочке, а второй — к противоположному полюсу глазного молока. Электроретинограмму можно отводить, помещая один электрод поверхности роговой оболочки и вводя второй электрод в нос или прикладывая его к коже лица вблизи глаза.
Зависимость возникновения электроретинограммы от фотохимических процессов в палочках и колбочках доказывается тем, что электрические реакции глаза возникают в онтогенезе одновременно с развитием с чувствительных рецепторов — палочек и колбочек — и появлением в зрительных пигментов. Содержание последних коррелирует с амплитудой волн электроретинограммы. В настоящее время еще неясно, каким образом расщепление зрительных пигментов вызывает изменение клеточных мембран, приводящее к возникновению электрической разности потенциалов. По-видимому, между фотохимическими и электрическими процессами существует какое-то промежуточное звено типа ферментативной реакции.
На электроретинограмме большинства животных, регистрируемой при освещении глаза в течение 1—2 секунд, различают несколько характерных волн (рис. 216). Первая волна — волна а — представляет собой небольшое по амплитуде электроотрицательное колебание, свидетельствующее об уменьшении потенциала покоя.
 |
Оно переходит в быстро нарастающую и медленно убывающую электроположительную волну в, имеющую значительно большую амплитуду. На фоне спада волны в нередко наблюдается медленная электроположительная волна с. В момент прекращения светового раздражения появляется еще одна электроположительная волна d. Рис. 216. Электрорстинограмма (по Р. Граниту). |
Электрорстинограмма человека имеет аналогичную форму с тем лишь отличием, что на ней между волнами а и в отмечается кратковременная волна х.
Р. Гранит, подробно проанализировавший форму электроретинограммы при различных воздействиях, пришел к заключению, что характерные ее волны обусловлены суммированием трех различных компонент. По его мнению, компонента, выявляющаяся в форме волны с, связана с палочковым зрением. Поэтому эта волна отсутствует на электроретинограмме тех животных (например, черепах), в сетчатке которых нет палочек.
Связь между волной с и палочковым зрением доказывается еще и тем, что при освещении глаза красным светом, действующим только на колбочки, на электроретинограмме нет волны с.
Вопрос о том, в каких структурах сетчатки возникают электрические колебания, регистрируемые в форме электроретинограммы, еще не решен. Для его выяснения была использована методика микроэлектродного отведения потенциалов от разных слоев сетчатки. При этом были получены разноречивые данные. По мнению одних исследователей, характерные волны электроретинограммы возникают в биполярных клетках. По мнению других, волна а электроретинограммы связана с процессом, происходящим во внешних члениках фоторецепторов, волна в возникает в наружном ядерном слое, а волна с — в пигментном эпителии. Амплитуда волны в электроретинограммы увеличивается пропорционально логарифму силы света; она возрастает также, если перед световым раздражением глаз длительное время находился в темноте. Волна d отражает реакции, возникающие в сетчатке при выключении света; чем длительнее глаз был подвергнут действию света, тем больше амплитуда волны d в момент прекращения светового раздражения.
Потенциалы действия в зрительном нерве. Медленные колебания электрических потенциалов при световом раздражении, регистрируемые в виде электроретинограммы, сопровождаются возникновением потенциалов действия в ганглиозных клетках сетчатки, от которых отходят волокна зрительного нерва. Впервые потенциалы действия в этом нерве были зарегистрированы в 1927 г. в опытах на морском угре Э. Эдрианом и Б. Метьюсом. Выбор объекта объяснялся тем, что у угря зрительный нерв состоит из небольшого числа относительно длинных волокон. Опыты показали, что в отсутствие световых раздражений — в темноте — потенцилов действия не было или же они были редкими.
Через 0,1—0,5 секунд после освещения глаза появлялась частая импульсная активность, причем в первый момент частота потенциалов была очень велика, а затем, несмотря на продолжающееся световое раздражение, она уменьшалась (рис.217). Сразу по прекращении действия света возникала вновь кратковременная вспышка импульсов в зрительном нерве. Связь между изменением мембранного потенциала в фоторецепторе и потоком импульсов в афферентном нервном волокне изучена Хартлайном в экспериментах на глазу мечехвоста (Limulus). Глаз этого животного состоит из отдельных чувствительных к свету образований — омматидиев в каждом из которых расположена нервная клетка, дающая начало в нервному волокну.
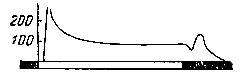 |
Риc. 217. Изменение частоты импульсов в зрительном нерве при освещении глаза. В нижнем части рисунка отмечено время освещения глаза (по Э. Эндриану и Б. Меттюсу). |
Хартлайн регистрировал микроэлектродом электрические потенциалы, возникающие в омматидии и в нервном волокне. Освещение глаза вызывает медленное электрическое колебание — рецепторный потенциал—равное 50 мв, вслед за которым появляются частые ритмические импульсы в нервном волокне (рис. 218). Чем выше иитенсивность светового раздражения, тем больше величина рецепторного потенциала и частота импульсов в нерве. При длительном освещении глаза рецепторный потенциал и частота импульсов в нервном волокне уменьшаются.
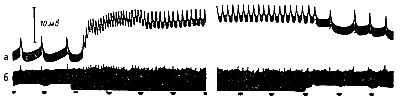 |
Рис. 218. Электрическая реакция глаза Limnlis на освещение (по Хартлайну). а — микроэлектродная регистрация электрических потенциалов одиночного омматидия, б — одновременная запись электрической активности нервных волокон, идущих от омматидия. |
В дальнейшем была зарегистрирована электрическая активность отдельных волокон зрительного нерва и у позвоночных животных. При этом были обнаружены три различные группы волокон. В первой группе волокон частая импульсная активность возникает лишь в начале светового раздражения и быстро затухает, во второй группе волокон частые импульсы появляются в момент прекращения действия света на глаз, а в третьей группе — и в начале и в момент прекращения светового раздражения (см. рис. 191). Очевидно, что первая группа волокон отходит от рецепторов, реагирующих на включение, вторая группа волокон связана с рецепторами, реагирующими на выключение, а третья группа волокон проводит импульсы от рецепторов, реагирующих и на включение и на выключении светового раздражения.
Отведение потенциалов действия от одиночного нервного волокна в сочетании с методикой точечного раздражения узким пучком световых лучей (диаметром около 0,1 мм) позволило установить площадь сетчатки, занимаемую фоторецепторами, стимуляция которых вызывает возбуждение одной и той же ганглиозной клетки. Эта площадь сетчатки представляет собой рецептивное поле данной клетки. Диаметр его равен примерно около 1 мм. Таким образом, одна ганглиозная клетка через посредство многих биполярных и горизонтальных нейронов (см. рис. 212) связана с тысячами фоторецепторов. Это соответствует гистологическим данным о структуре сетчатки и тому, что на 130 млн. палочек и колбочек приходится всего около 1 млн. нервных волокон зрительного нерва. Середина рецептивного поля (диаметром около 0,2 мм в области центральной ямки и около 0,6 мм в периферических частях сетчатки) обладает максимальной чувствительностью. Края рецептивного поля оной и той же ганглиозной клетки обладают меньшей чувствительностью.
Если 2 узких луча света — оба допороговой интенсивности — действуют в пределах одного рецептивного поля, то происходит суммация рецепторных потенциалов и в ганглиозной клетке возникает импульсации, регистрируемая в нервном волокне зрительного нерва.
Если же 2 узких луча света — сворхпороговой интенсивности — действуют на
разные рецептивные поля, относящиеся к разным ганглиозным клеткам сетчатки, то
наблюдаются явления торможения; возбуждение одного рецептивного поля повышает
порог раздражения другого рецептивного поля. Таким образом, нейронам сетчатки
свойственны те же самые явления (суммация, торможение), которые характерны для
нервных центров. Это дало основание считать нейроны сетчатки вынесенной на
периферию частью центральной нервной системы.
