Более 100 лет назад И.М. Сеченов экспериментами на лягушках показал, что
раздражение зрительных бугров кристаллами поваренной соли вызывает угнетение
рефлексов спинного мозга. На основание своих опытов ученый сделал два
кардинальных открытия. Одно из них — открытие процесса торможения — было оценено сразу. Второе — открытие
ретикуло-спинальных влияний — получило широкое признание лишь в последние 20
лет, после работ Г. Мэгуна и его сотрудников.
Г. Мэгун и Р. Рейнис показали, что электрическое раздражение участков
ретикулярной формации продолговатого мозга животного тормозит спинномозговые
рефлексы (рис. 232, А), а после децеребрации животного снижает
ригидность мускулатуры. При слабых раздражениях ретикулярной формации одной
стороны торможение охватывает только нейроны спинного мозга на этой же стороне.
При более сильных раздражениях тормозятся нейроны обеих половин спинного мозга;
торможо; обнаруживается при исследовании как сгибатсльпых, так и разгибательных
рефлексов. Подобные эффекты возникают лишь при раздражении вентромедиальной
части продолговатого мозга. Раздражение других участков ствола не приводит к
подобным явлениям.
Перерезки спинномозговых проводящих путей позволили установить ход волокон,
по которым распространяются импульсы от нейронов ретикулярной формации,
вызывающих угнетение рефлексов спинного мозга. Эти волокна действуют на клетки
Реншоу и усиливают их тормозящий эффект на мотонейроны. Кроме того, импульсы,
приходящие из ретикулярной формации, могут затормаживать и непосредственно
активность мотонейронов.
После кратковременного раздражения участков ретикулярной формации возникает
облегчение рефлекторной деятельности спинного мозга. На этом основании высказано
предположение, что в ретикулярной формации имеются нейроны, оказывающие
активирующее влияние на нервные клетки спинного мозга. Эксперименты с
раздражением различных участков мозгового ствола подтвердили это предположение.
Оказалось, что в промежуточном мозгу — в гипоталамусе, в сером веществе покрышки
среднего мозга и варолиева моста и в продолговатом мозгу к периферии от тех
участков, раздражение которых оказывает тормозящее влияние, расположены нейроны
ретикулярном формации, активирующие рефлекторную функцию спинного мозга
(рис. 232, Б).
 |
|
Раздражение этих участков усиливает спинномозговые рефлексы и
сокращения скелетной мускулатуры, вызванные раздражением коры больших
полушарий (рис. 232, Б).
Рис. 232. Эффекты раздражении разных участков ретикулярной
формации; тормозящего (А) и активирующего спинной мозг (Б) (по Г. Мэгуну).
Ни рисунке А — записи сокращений мышц при сгибатсльном (а), коленном (б) и
мигательном (в) рефлексах. На рисунке Б — записи сокращений мышц в ответ
на раздражение коры больших полушарий (а) и при коленном рефлексе (б).
Внизу на обоих рисунках — отметка раздражении ретикулярной
формации. |
Прослеживание путей, по которым проводятся импульсы, активирующие нейроны
спинного мозга, показало, что ими являются волокна ретикуло-спинального тракта.
При этом выяснено, что волокна, активирующие и тормозящие, различны.
Активирующие волокна ретикулярной формации оканчиваются на вставочных нейронах
рефлокторных дуг. Не исключено, что облегчение спинномозговых рефлексов под
влиянием ретикулярной формации происходит вследствие подавления тормозящих
разрядов клеток Реншоу, что приводит к повышению возбудимости мотонейронов.
Ретикулярная формация оказывает влияние не только на рефлекторные движения
(физические рефлексы), но и на тонус скелетной мускулатуры (тонические
рефлексы).
Устранение активирующего и тормозящего действия ретикулярной формации при
высокой перерезке спинного мозга является, по-видимому, одной из причин
возникновения спинального шока и развивающейся позже гиперрефлексии.
Экспериментально доказана роль ретикулярной формации в возникновении
децеребрационной ригидности. Перерезка мозгового ствола выше продолговатого
мозга препятствуот притоку импульсов, понижающих активность тех нейронов
бульбарной ретикулярной формации, которые оказывают тормозное влияние на центры
спинального мозга (см. рис. 233). Вместе с тем нейроны, оказывающие
активирующее влияние, находящиеся в области покрышки варолиева моста, продолжают
получать афферентные импульсы от вестибулярных ядер и от спинного мозга. Таким
образом нарушается баланс между тормозными и облегчающими импульсами
ретикулярной формации в сторону преобладания активирующих импульсов. В
результате возникает резкое повышение тонуса скелетной, особенно разгибательной,
мускулатуры. При разрушении участков активирующей ретикулярной формации и
области покрышки варолиева моста децеребрационная ригидность не развивается.
 |
|
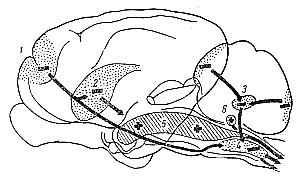
Рис. 233. Схематическое изображение мозга кошки, на котором указаны
обличающие (+) и тормозящие (—) зоны ретикулярной формации ствола мозга и
его связи с корой, подкорковыми структурами и мозжечком (по Г. Мэгуну).
Тормозящие пути: 1 — кортико-ретикулярный путь; 2 — каудато-спинальный
путь; 3— мозжечково-ретикулярый путь; 4— ретикуло-спинальный путь.
Облегчающие пути: 5 — ретикуло-спинальный путь; 6— вестиобуло -спинальный
путь. |
При повреждениях некоторых участков среднего мозга возникает ригидность
преимущественно сгибательной мускулатуры. При очень ограниченных повреждениях
ретикулярной формации среднего мозга может быть вызвана ригидность только одной
конечности.
Механизм влияния ретикулярной формации на мышечный тонус с известен благодаря
работам Р. Гданита, который показал, что она изменяет
активность гамма-мотонейронов спинного мозга. Последние
иннервируют мышечные волокна периферических частей мышечных веретен (стр. 456).
Свое название они получили потому, что их аксоны, так называемые
гамма-афференты, представляют собой тонкие волокна, относящиеся к типу Аγ,
обладающие значительно меньшей скоростью проведения, моторные
волокна скелетных мышц.
Как уже говорилось (стр.457), гамма-эфференты, вызывая сокращение мышечных
элементов веретен, обусловливают их натяжение и следствие этого усиление
афферентной импульсации рецепторов ядерной сумки веретен.
Афферентные импульсы от веретен, постоянно поступая к спинному мозгу,
возбуждают альфа-мотонейроны, что и является причиной тонуса мышц. Роль
афферентных импульсов в мышечном тонусе демонстрируется тем, что перерезка
задних корешков спинного мозга приводит к исчезновению мышечного
тонуса. В свою очередь поток афферентных импульсов от веретен регулируется
гамма-мотонейронами.
Таким образом, имеются сложные взаимоотношения и обратные связи между
нейронами спинного мозга и скелетной мускулатурой. Эти взаимоотношения
регулируются ретикулярной формацией, которая посредством своего влияния на
гамма-мотонейроны изменяет поток афферентных импульсов от веретен и тем самым
влияет на мышечный тонус.
Регуляция мышечного тонуса осуществляется покрышкой среднего мозга по
двум ретикуло-спинальным путям — быстропроводящему медленнопроводящему. По
первому проводятся импульсы, контролирующие быстрые движения; импульсы,
проходящие по второму пути, контролируют медленные тонические сокращения.
Ретикуло-спинальные механизмы находятся под постоянным контролем мозжечка и
коры больших полушарий головного мозга.


