

Вестибулярный аппарат и рецепция положения и движения тела
Рецепторы вестибулярного аппарата доставляют информацию о положении и движении тела в пространстве. Поступающие в центральную нервную систему импульсы от рецепторов вестибулярного аппарата обусловливают возникновение рефлексов, необходимых для сохранения равновесия тела. В результате этих рефлексов происходят сложнокоординированные тонические сокращения скелетной мускулатуры, с помощью которых положение тела выравнивается и сохраняется равновесие.
В пирамиде височной кости находится внутреннее ухо, или лабиринт, который состоит из улитки и вестибулярного аппарата. Последний включает в себя преддверие (vestibulnm) и три полукружных канала (саnales semicirculares). Полукружные каналы, представляющие собой узкие ходы правильной формы, расположены не совсем точно в трех взаимно перпендикулярных плоскостях. Верхний, или передний, канал лежит во фронтальной, задний — в сагиттальной, а наружный— в горизонтальной плоскости. Один конец каждого канала колобообразно расширен и называется ампулой.
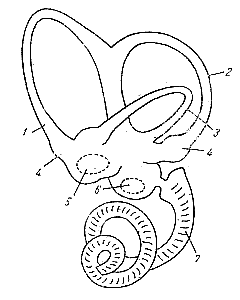 |
В костном лабиринте имеется почти точно повторяющий его форму перепончатый лабиринт. Последний в области предверия делится на два мешочка: один - sacculus — помещается в сферическом вдавлении кости и лежит ближе к улитке, другой—utriculus — лежит в имеющем эллиптическую форму вдавлении кости и расположен ближе к полукружным каналам (рис. 196). Рис. 196. Схема левого лабиринта млекопитающего. 1, 2 и 3 — полукружные каналы; 4 — ампулы каналов; 5, 6 — преддверие (vestibulum), которое разделяется на два мешочка: utriculus (3) и sacculus (б); пунктиром отмечены «слуховые» пятка — macula utrlculae и macula sacculac; 7 — улитка. Между костью и перепончатым лабиринтом имеется небольшое пространство, заполненное жидкостью, — перилимфой. Жидкость внутри перепончатого лабиринта называется эндолимфой; вязкость в 2—3 раза больше, чем воды. В мешочках преддверия — sacculus utriculus — находится так называемый отолитовый аппарат, имеющий сложную структуру. |
В каждом мешочке имеются возвышения, называемые пятнами — macula sacc и macula utriculi, представляющие собой скопления рецепторных клеток. Одна половина каждой такой клетки имеет цилиндрическую форму, другая — сужена и снабжена одним подвижным волоском и 60-80 склеенными неподвижными волосками (рис. 197). Волоски рецепторных клеток проникают в сетчатую, похожую на войлок ткань, имеющую студенистую консистенцию и покрывающую maculaе.
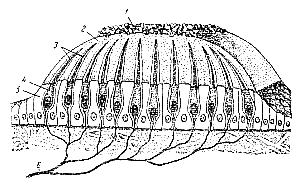 |
В петлях этой студенистой массы находится большое количество микроскопических образований, состоящие из органического соединения фосфорно-углексилого кальция и имеющих форму продолговатых шестиугольников. Эти образования называются отолитами. Отолитовый слой, или отолитовая мембрана, тяжелее остальной ткани и может оказывать механическое давление на волоски рецепторных клеток. Рис. 197. Схема строения отолитового аппарата (из Кольба). 1 — отолиты; а — отолитовая мембрана; 3 — полоски рецепторных клеток; 4 — рецепторные клетки; 5 — опорные клетки; б — нервные волокна. |
В перепончатых полукружных каналах рецепторные клетки имеются только в ампулах, где они образуют скопления, получившие название cristae ampularis. Эти рецепторпые клетки имеют длинные волоски. При движении эндолимфы каналов, происходящем при вращении тела; может происходить изгиб волосков рецепторных клеток.
К рецепторным клеткам вестибулярного аппарата подходят нервные волокна, являющиеся отростками биполярных клеток, образующих ganglion scarpae. Между реценторной клеткой в дендритом биполярного нейрона имеется синапс, передача возбуждения в котором осуществляется посредством медиатора — ацетилхолина. Вторые отростки биполярных нейронов по выходе из ganglion scarpae образуют вестибулярную ветвь VIII пары черепномозговых нервов.
Адекватными раздражителями вестибулярного аппарата является ускоренное или замедленное вращательное или прямолинейное движение тела. Раздражение рецепторов вестибулярного аппарата происходит лишь под влиянием изменения скорости движения. Равномерное движение без ускорения или замедления не вызывает раздражения.
Электрофизиологические исследования подтвердили эти давно установленные факты. По афферентным нервным волокнам, отходящим от вестибулярного аппарата, постоянно поступают в центральную нервную систему редкие импульсы, даже в условиях покоя. Резкое возрастание частоты импульсов наблюдается в начале и в конце вращательного движения. Частые залпы импульсов длятся при этом около 25 секунд. Если вращательное движение равномерно, то импульсация в афферентных нервных волокнах, отходящих от полукружных каналов, отсутствует. В зависимости от характера движения может возникнуть преимущественное раздражение отолитовых рецепторов или же рецепторов полукружных каналов.
Раздражителями отолитового аппарата являются ускоряющееся или замедляющееся прямолинейное движение тела, а также тряска, качка и наклон тела или головы в сторону, вызывающие изменения давления отолитовой мембраны на волоски рецепторных клеток.
Раздражение рецепторов отолитового аппарата, согласно одной теории— теории Брейера, происходит, когда отолитовая мембрана в силу собственной тяжести или под влиянием движении головы отодвигается внутрь преддверия и натягивает волоски рецепторных клеток. Согласно этой теории, реценторные клетки подвергаются раздражению в том случае, когда отолитовая мембрана под влиянием тех же причин надавлиет на их волоски.
Рецепторы отолитового аппарата при прямолинейном движении дают возможность воспринять ускорение, равное 2—20 см/сек2 (порог различения ускорения). Порог различения наклона головы в сторону составляет около 1°, вперед и назад — около 1,5—2°.
При сопутствующих раздражениях можно наблюдать значительное повышение этого порога, т. е. ухудшение различения наклона, например, вибрации, имеющиеся в самолетах, повышают этот порог до 5° при наклонах вперед и назад и до 10° при наклонах в сторону.
Раздражителем рецепторов полукружных каналов является ускоренное или замедленное вращательное движение в какой-либо плоскости. В зависимости от характера вращательного центробежного ускора или замедления происходит неодинаковое раздражение рецепторов разных полукружных каналов: рецепторы канала, расположенного в oдной плоскости с направлением вращения, раздражаются с большей силой, чем другие.
Порог различения вращения равен в среднем 2—3°/сек2 углового ускорения.
Раздражителем рецепторов полукружных каналов является движение эндолимфы. Это доказано опытами Эвальда: посредством тонкой операции он вводил животному в один из полукружных каналов «пневматический молоточек», который мог производить перемещение эндолимфы, когда его приводили в движение. Эксперименты показали, что при этом возникают такие же рефлекторные движения животного, как и при вращении в соответствующую сторону.
