

Связь спинного мозга с вышележащими отделами центральной нервной системы (мозговым стволом, мозжечком и большими полушарием осуществляется посредством восходящих и нисходящих проводящих путей. По восходящим путям передается информация, получаемая рецепторами.
Импульсы от проприорецепторов мышц, сухожилий и связок проходят в вышележащие отделы центральной нервной системы частью по волокнам пучковГолля и Бурдаха, находящимся в задних столбах спинного мозга, частью по волокнам спино-мозжечковых путей Говерса и Флексига, расположенных в боковых столбах. Пучки Голля и Бурдаха образованы отростками рецепторных нейронов, тела которых находятся в спинномозговых ганглиях (рис. 227).
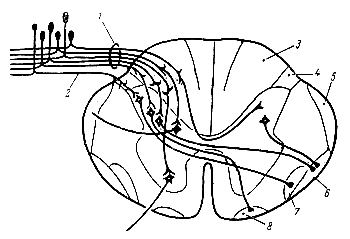 |
Рис. 227. Схематическое изображение хода пол окон задних корешков в спинном мозгу (по Ренсону и Кларку). 1 — толстые волокна заднего корешка; 2 — тонкие волокна заднего корешка; 3 — задние столбы; 4 — заднелатеральный канатик (тракт Лиссауера); 5 — дорсальный спино-мозжечковый тракт; 6 — вентральный спино-мозжечковый тракт; 7 — латеральный спино-таламический тракт; 8 — вентральный спино-таламический тракт. |
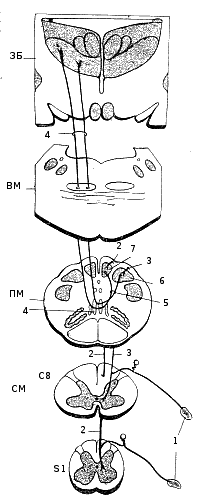 |
Эти отростки, войдя в спинной мозг, идут в восходящем направлении, отдавая короткие ветви к серому веществу нескольких выше и ниже расположенных сегментов спипного мозга. Эти ветви образуют синапсы на промежуточных и эффекторных нейронах, входящих в состав спинномозговых рефлекторных дуг. Пучки Голля и Бурдаха оканчиваются в ядрах продолговатого мозга, откуда начинается второй нейрон афферентного пути, направляющийся после перекреста к таламусу; здесь расположен третий нейрон, отростки которого проводят афферентные импульсы к коре больших полушарий (рис. 228). За исключением тех волокон, которые входят в состав пучков Голля и Бурдаха и идут, не прерываясь, в продолговатый мозг, все остальные афферентные нервные волокна задних корешков вступают в серое вещество спинного мозга и здесь прерываются, т. е. образуют синапсы на различных нервных клетках. От так называемых столбовых, или кларковых, клеток заднего рога и отчасти от спайковых, или комиссуральных, клеток спинного мозга берут начало нервные волокна пучков Говерса и Флексига. Нарушение проведения афферентных импульсов по спино-мозжечковым путям влечет за собой расстройство сложных движений, при которых наблюдаются нарушения мышечного тонуса и явления атаксии, как и при поражениях мозжечка. Рис. 228. Схема проводящих путей задних столбов спинного мозга. 1 — тактильные рецепторы кожи; 2 — нежный пучок Голля (fasciculus gracilis); 3 — клиновидный пучок Бурдаха (fasciculus cuneatus); 4 — медиальная петля (lemniscus medians); 5 — перекрест медиальной петли; 6 — ядро Бурдаха в продолговатом мозгу; 7 — ядро Голля в продолговатом мозгу; СМ — спинной мозг (сегменты С8 и S1); ПМ — продолговатый мозг; ВМ — варолиев мост; ЗБ — зрительные бугры (видны ядра, особенно заднее вентральное, где заканчиваются волокна медиальной петли). |
Импульсы от проприорецепторов распространяются по обладающим высокой скоростью проведения (до 140 м/сек) толстым миелиновым волокнам группы Аα, образующим спино-мозжечковые пути, и по более медленно проводящим (до 70 м/сек) волокнам пучков Голля и Бурдаха. Большая скорость проведения импульсов от рецепторов мышц суставов и сухожилий, очевидно, связана с важностью для организма быстрого получения информации о характере выполняемого двигательного акта, что обеспечивает непрерывный его контроль.
Импульсы от болевых и температурных рецепторов поступают к клеткам задних рогов спинного мозга; отсюда начинается второй нейрон афферентного пути. Отростки этого нейрона на уровне этого же сегмента, где расположено тело нервной клетки, переходят на противоположную сторону, вступают в белое вещество боковых столбов и в составе латерального спино-таламического пути (см. рис. 227) идут к зрительному бугру, где начинается третий нейрон, проводящий импульсы к коре больших полушарий. Импульсы от болевых и температурных рецепторов частично проводятся и по волокнам, направляйся кверху по задним рогам серого вещества спинного мозга. Проводники болевой и температурной чувствительности представляют собой тонкие миелиновые волокна группы АΔ и безмиелиновые волокна, отличающиеся малой скоростью проведения.
При некоторых поражениях спинного мозга могут наблюдаться расстройства только болевой или только температурной чувствительности. Более того, может быть нарушена чувствительность только к теплу или только к холоду. Это доказывает, что импульсация от соответствующих рецепторов проводится в спинном мозгу по нервным волокнам.
Импульсы от тактильных рецепторов кожи поступают к клеткам задних рогов, отростки которых восходят по серому веществу на несколько сегментов, переходят на противоположную сторону спинного мозга, вступают в белое вещество и в вентрального спино-таламического пути несут импульсь к ядрам зрительных бугров, где находится третий нейрон, передающий получаемую им информацию коре больших полушарий. Импульсы от кожных рецепторов прикосновения и давления частично проходят также по пучкам Голля и Бурдаха.
Имеются существенные различия в характере информации, доставляемой волоканми пучков Голля и Бурдаха и волокнами спино-таламических путей, а также в скорости распространения импульсов по тем и другим. По восходящим путям задних столбов передаются импульсы от рецепторов прикосновения, обеспечивающие возможность точной локализации места раздражения. Волокна этих путей проводят также импульсы большой частоты, возникающие при действии вибрации на рецепторы. Здесь же проводятся импульсы от рецепторов давления, дающие возможность точного определения интенсивности раздражения. По спино-таламическим путям проводятся импульсы от рецепторов прикосновения, давления, а также от температурных и болевых рецепторов, не обеспечивающие точной дифференцировки локализации и интенсивности раздражения.
Волокна, проходящие в пучках Голля и Бурдаха, передающие более дифференцированную информацию о действующих раздражениях, проводят импульсы с большей скоростью, причем частота этих импульсов может меняться в значительных пределах. Волокна спино-таламических путей обладают малой скоростью проведения; при разной силе раздражения частота импульсов, проходящих в них, мало меняется.
Импульсы, которые проводятся по афферентным путям, генерируют, как правило, возбуждающий постсинаптический потенциал, достаточно сильный для того, чтобы вызвать возникновение распространяющегося импульса в следующем нейроне восходящего афферентного пути. Однако импульсы, переходящие с одного нейрона на другой, могут затормаживаться, если в данный момент центральная нервная система получает по другим афферентным проводникам какую-либо более важную для организма информацию.
По нисходящим путям спинного мозга поступают к нему импульсы от вышележащих эффекторных центров. Получая импульсы по нисходящим путям от центров головного мозга и передавая эти импульсы к рабочим органам, спинной мозг выполняет проводниково-исполнительскую роль.
По кортикоспинальным, или пирамидным, путям, проходящим в передних боковых столбах спинного мозга, к нему приходят импульсы непосредствено от крупных пирамидных клеток коры больших полушарий. Волокна пирамидных путей образуют синапсы на промежуточных и моторных нейронах (прямая связь пирамидных нейронов с мотонейронами имеется только у человека и обезьян). В составе кортикоспинальных путей имеется около миллиона нервных волокон, среди которых около 3%составляют толстые волокна диаметром 16 мк, относящиеся к типу Аα и обладают большой скоростью проведения (до 120—140 м/сек). Эти волокна представляют собой отростки крупных пирамидных клеток коры. Остальные волокна имеют диаметр около 4 мк и обладают гораздо меньшей скоростью проведения. Значительное количество этих волокон проводит импульсы к спинальным нейронам вегетативной нервной системы.
Кортикоспинальные пути боковых столбов перекрещиваются на уровне нижней трети продолговатого мозга. Кортикоспинальные пути передних столбов (так называемые прямые пирамидные пути) не перекрещиваются в продолговатом мозгу; они переходят на противоположную сторону вблизи того сегмента, где заканчиваются. В связи с этим перекрестом кортикоспинальных путей нарушения моторных центров одного полушария вызывают паралич мускулатуры противоположной стороны тела.
Через некоторое время после повреждения пирамидных нейронов или идущих от них нервных волокон кортикоспинального тракта возникают некоторые патологические рефлексы. Типичным симптомом поражения пирамидных путей является извращенный кожно-подошвенный рефлекс Бабинского. Он проявляется в том, что штриховое раздражение подошвенной поверхности стопы вызывает разгибание большого пальца и веерообразное расхождение остальных пальцев ноги; такой рефлекс получается также и у новорожденных, у которых пирамидные пути еще не закончили своего развития У здоровых взрослых людей штриховое раздражение кожи подошвы вызывает рефлекторное сгибание пальцев.
В синапсах, образованных волокнами кортикоспинального тракта, могут возникать как возбуждающие, так и тормозящие постсинаптические потенциалы. В результате может возникать возбуждение или торможение мотонейронов.
Аксоны пирамидных клеток, образующие кортикоспинальные пути, отдают коллатерали, которые заканчиваются в ядрах полосатого тела, гипоталамуса, и красном ядре, в мозжечке, в ретикулярной формации мозгового ствола. От всех перечисленных ядер импульсы по нисходящим путям, называемым экстракортикоспинальными, или экстрапирамидными, поступают к вставочным нейронам спинного мозга. Главными из этих нисходящих путей являются ретикуло-спинальный, рубро-спинальный, текто-спинальный и вестибуло-спинальный тракты. По рубро-спинальному тракту (пучку Монакова) к спинному мозгу поступают импульсы от мозжечка, четверохолмия и подкорковых центров. Импульсы, проходящие по этому пути, имеют значение в координации движении и регуляции тонуса мышц.
Вестибуло-спинальный тракт идет от вестибулярных ядер в продолговатом мозгу к клеткам переднего рога. Импульсы, приходящие по этому пути, обеспечивают осуществление тонических рефлексов положения тела. Ретикуло-спинальные пути передают активирующее и тормозящее влияния ретикулярной формации на нейроны спинного мозга. Они оказывают влияние- как на моторные, так и на промежуточные нейроны. Кроме всех этих длинных нисходящих путей (в белом веществе спинного мозга), имеются еще и короткие пути, связывающие вышележащие сегменты с нижележащими.
