

Описанный механизм стойкой деполяризации постсинаптической мембраны при очень частом следовании друг за другом нервных импульсов лежит в основе открытого Н. Е. Введенским явления пессимума. Это явление часто называют «торможением Введенского». Сущность его состоит в следующем. Высота тетанического сокращения скелетной мышцы в ответ на ритмические раздражения нерва возрастает с увеличением раздражения.
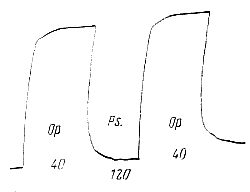 |
При некоторой оптимальной (от слова "optimum" - наилучший) частоте раздражений тетанус достигает наибольшей величины. Если продолжать и дальше повышать частоту стимуляции нерва, то вместо увеличения тетаническое сокращение мышцы начинает резко и при некоторой большой, пессимальной (от слова «pessimum» — наихудший) частоте раздражения нерва мышца, несмотря на продолжающееся раздражение, почти полностью расслабляется. Уменьшение частоты стимуляции тотчас же приводит к восстановлению исходного высокого уровня тетанического сокращения (рис.163). Рис. 163. Оптимум (Ор.) и пессимум (Ps.) в нервно-мышечном препарате. Цифры на рисунке обозначают частоту раздражения. |
В нервно-мышечном препарате лягушки пессимум возникает при частотах раздражения в пределах 50—100 стимулов в секунду, т.е. при значительно меньших частотах, чем те, которые может воспроизводить нерв (порядка 500—1000) или мышца (порядка 200—300).
Лабильность синапсов намного ниже лабильности нервных и мышечных волокон. Это объясняют тем, что передача возбуждения через синапс связана с процессами освобождения и последующего ферментативного расщепления медиатора, т. е. процесса, происходящего относительно медленно.
На рис. 164 приведена запись изменений мембранного потенциала мышечного волокна в области концевой пластинки при раздражении нерва ритмическими импульсами оптимальной и пессимальной частоты. При редком— оптимальном — ритме каждый следующпй импульс попадает в фазу убывания постсинаптического потенциала, вызванного предыдущим импульсом, и в мышечном волокне возникают полноценные потенциалы действия.
 |
При частом же раздражении постсинаптические потенциалы суммируются, что приводит к стойкой деполяризации постсинаптической раны и развитию блока проведения. На роль ацетилхолина в явлении пессимума указывает тот факт, что яды, парализующие холинэстеразу и тем самым способствующие накоплению в области синапса ацетилхолина, водят к возникновению пессимума (А. Г. Гинецинский). Рис. 164. Стойкая деполяризация постсинаптической мембраны
мышечного волокна при песссимальном раздражении двигательного нерва (по Н.
М. Шамариной). |
Следует, однако, подчеркнуть, что описанный механизм блокирования нервно-мышечного соединения при частом ритме раздражения нерва не является единственным. Выяснилось, что в тех случаях, когда частота стимулов очень высока, проведение возбуждения с нерва на мышцу может быть блокировано еще на пути к синапсу, в тонких пресинаптических разветвлениях нервных волокон (пресинаптических терминалях), обладающих более низкой лабильностью, чем толстые нервные волокна до их концевых разветвлений.
Нарушение проведения в этих тонких нервных волокнах приводит к прекращению поступления нервных импульсов к нервному окончанию и тем самым к прекращению выделения ацетилхолина. В зтом случае вместо стойкой деполяризации постсинаптической мембраны обнаруживается значительное ослабление или даже полное выпадение постсинаптических потенциалов при неизменённом уровне потенциала покоя мышечного волокна.
