

Потенциал действия, как привило, сопровождается так называемыми следовыми потенциалами. Они были впервые зарегистрированы Д. С. Воронцовым (1926), а в дальнейшем подробно изучены Дж. Эрлангером и Г. Гассером и др.
Различают отрицательные и положительные следовые потенциалы. Амплитуда как тех, так и других не превышает нескольких милливольт, а длительность варьирует от нескольких миллисекунд до нескольких десятков или даже сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, медленно развивающимися в нервных и мышечных волокнах после окончания возбуждения.
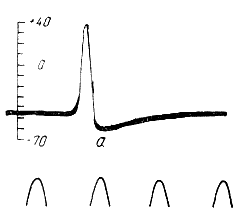
Взаимоотношения между потенциалом действия и следовым отрицательным потенциалом могут быть рассмотрены на примере потенциала действия скелетного мышечного волокна. Из записи, приведенной на рис. 118, видно, что фаза реполяризации делится на две неравные по длительности части. Вначале реполяризация мембраны идет быстро, а затем замедляется и приостанавливается. Этому моменту и соответствует начало следового отрицательного потенциала (в). Мембрана в течение некоторого времени остается частично деполяризованной, лишь примерно через 15 мсек происходит полное восстановление мембранного потенциала до исходной величины — 85 мв. Следовой отрицательный потенциал часто называют следовой деполяризацией мембраны.
Следовой положительный потенциал выражается в усилении нормальной поляризации — гиперполяризации — мембраны. Он особенно хорошо выражен в безмякотных нервных волокнах. Так, в безмякотном гигантском аксоне кальмара нисходящая фаза потенциала действия непосредственно переходит в положительный следовой потенциал, амплитуда которого достигает примерно 15 мв. и лишь затем мембранный потенциал возвращается к исходному уровню покоя (рис. 119).
В миелинизированных нервных волокнах следовые изменения потенциала имеют более сложный характер: следовой отрицательный потенциал часто сменяется следовым положительным потенциалом, затем иногда развивается новая электроотрицательность и лишь после этого происходит полное восстановление потенциала покоя.
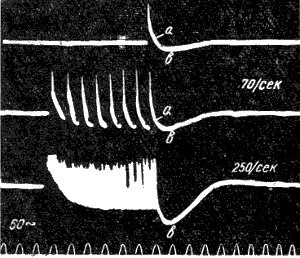
При ритмическом раздражении нерва следовые потенциалы суммируются, вследствие чего их амплитуда и длительность возрастают (рис. 120).
 |
 | |
| Рис. 119. Потенциал действия гигантского аксона кальмара, записанный с помощью внутриклеточного микроэлектрода (по Ходжкину и Хаксли). Выражен следовой положительный потенциал (а). В нижней части рисунка отметка времени — 500 колебании в 1 секунду. |
Рис. 120. Суммация следовых потенциалов в диафрагмальном нерве кошки при кратковременном его раздражении ритмическими стимулами. Восходящая часть потенциалов действия не видна. Записи начинаются с отрицательных следовых потенциалов (а), переходящих в положительные потенциалы (в). Верхняя кривая показывает ответ на одиночное раздражение. С увеличением частоты стимуляции следовой положительный потенциал резко возрастает (по Гассеру). |
