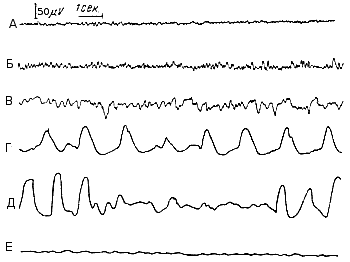
Ритмы электроэнцефалограммы - по частоте, амплитуде и физиологическим характеристикам электрических колебаний в электроэнцефалограмме различают четыре основных типа (рис. 243).
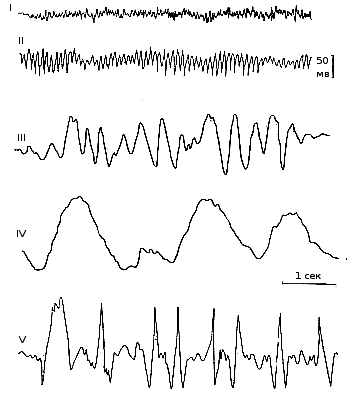 |
Альфа-ритм — это ритмические колебания потенциала почти синусоидальной формы, частотой 8—13 в секунду, с амплитудой до 50 мкв. Альфа-ритм отчетливо выражен, если испытуемый человек находится в условиях физического и умственного покоя — лежа или сидя в удобном кресле, с расслабленной мускулатурой и закрытыми глазами, при отсутствии внешних раздражений. Многие исследователи считают, что существует две области коры, в которых альфа-ритм имеет наибольшую амплитуду и характеризуется большим постоянством: одна из них находится в затылочной, вторая — в теменной доле. Затылочный альфа-ритм возникает в зрительной зоне коры; он, как правило, отсутствует или слабо выражен у слепых. Теменной альфаритм называется роландическим, так как он связан с активностью роландической области, в которой находится корковый конец проприорецептивного (двигательного) анализатора. Колебания, похожие на альфа-ритм человека, регистрируются в сходных условиях у лабораторных животных и называются альфаподобными ритмами. Рис. 243. Основные ритмы электроэнцефалограммы (схематизировано). I — бета-ритм; II — альфа-ритм; III — тета-ритм; IV — дельта-ритм; V — судорожные разряды. |
Бета-ритм характеризуется частотами колебаний выше 13 в секунду и амплитудой до 20-25 мкв. Этот ритм наиболее выражен в лобных и несколько меньше в теменных отделах коры. В затылочной области коры альфа-ритм быстро сменяется бета-ритмом при нанесении различных, особенно световых, раздражений, при умственной работе, например при решении арифметической задачи, эмоциональном возбуждении и т. п. Чем больше напряжение внимания при умственной деятельности или чем сильнее раздражение, действующее на рецепторы, тем быстрее альфа-ритм сменяется бета-ритмом. Роландический ритм также сменяется бета-ритмом при различных раздражениях, но особенно резко на него влияют проприорецептивные раздражения, возникающие при движении конечностей.
Тета-ритм представляет собой колебания потенциалов, частотой 4-8 в секунду, с амплитудой 100-150 мкв. Он наблюдается во время сна и при различных патологических условиях: при гипоксии и умеренно глубоком наркозе.
Дельта-ритм характеризуется медленными колебаниями потенциалов частотой 0,5-3,5 в секунду, с амплитудой до 250-300 мкв. Дельта-волны регистрируются во время глубокого сна, при глубоком наркозе, гипоксии и различных патологических процессах в коре больших и полушарий.
Вопрос о происхождении волн электроэнцефалограммы в настоящее время еще не решен полностью. Исследования с помощью внутриклеточных или внеклеточных микроэлектродов показывают, что нейроны коры, так же как клетки других отделов центральной нервной системы, способны генерировать потенциал действия при их раздражении или поступлении к ним импульсов от других нервных клеток. Вкоре больших полушарий в многочисленных синапсах, кроме того, возникают предшествующие потенциалам действия и значительно медленнее протекающие постсинаптические (возбуждающие и тормозные) потенциалы.
Предполагалось, что медленные волны электроэнцефалограммы представляют собой алгебраическую сумму потенциалов действия множества асинхронно работающих одиночных нейронов. Однако эта точка зрения, выдвинутая Э. Эдрианом, в настоящее время оставлена, так как доказано, что между импульсной активностью одиночных нейронов и волнами электроэнцефалограммы отсутствует какая-либо связь (рис. 244). При некоторых же воздействиях эти два вида активности могут полностью расходиться. Например, при эфирном наркозе клетки коры утрачивают способность к генерации потенциалов действия, между тем как медленные колебания потенциалов электроэнцефалограммы продолжают регистрироваться и оказываются усиленными.
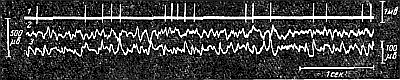 |
Рис. 244. Одновременная запись импульсной активности отдельного нейрона коры (микроэлектродное отведение) (1) и электроэнцефалограммы, отведенной из глубины коры (2) и с ее поверхности (3) у кошки (по В. Б. Головчинскому). |
По мнению большинства исследователей, происхождение волн электроэнцефалограммы связано с алгебраической суммацией постсинаптических потенциалов. Результаты их суммации будут различны в зависимости от прихода импульсов в кору больших полушарий из других отделов центральной нервной системы.
При синхронном возбуждении большой группы нервных клеток коры больших полушарии в результате суммации постсинаптических потенциалов на отводящих электродах регистрируются высокоамплитудные, медленные, альфа- или дельта-подобные волны электроэнцефалограммы. Такое состояние наблюдается при ограничении поступления афферентных импульсов в кору (при закрывании глаз, при пребывании в тихом и затемненном помещении), а также во время сна и наркоза.
Если же в кору поступают афферентные импульсы, то в различных клетках, находящихся под электродом, постинаптические потенциалы возникают пеодновременно и на электроэнцефалограмме регистрируются частые низкоамплитудные колебания потенциалов типа бета-ритма. Такие изменении происходят в электроэнцефалограмме при раздражении от сна и при бодрствовании. Переход от медленных ритмов к частым наблюдается в электроэнцефалограмме при раздражении ретикулярной формации ствола мозга. Это явление получило название реакции десинхронизации, или активации (Г. Джаспер). Одним из примеров возникновения такой реакции является смена альфаритма бета-ритмом при открывании глаз человеком (рис. 244А).
О ведущей роли афферентной импульсации в возникновении и педдержании ритмов электроэнцефалограммы свидетельствуют опыты с так называемой подрезкой коры. Если пересечь все нервные волокна, приходящие к небольшому участку коры из подкорковых ядер, но при этом сохранить его связи с сосудистым руслом (препарат «изолированной полоски коры»), то электрическая активность в этом участке полностью прекращается. Однако если этот участок подвергнуть прямому электрическому раздражению, в нем вновь появляется серия постепенно затухающих колебаний потенциалов.
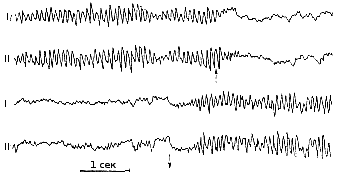 |
Рис. 244 А. Изменения электроэнцефалограммы затылочной области коры, показывающие переход от альфа-ритма к бета ритму при открывании глаз (стрелка вверх) и восстановление альфа-ритма при закрывании глаз (стрелка вниз). |